0 – A mo’ di introduzione
Per annunciare al modo che esistono piante a ovulo nudo (non contenuto in un ovario) bisogna sapere bene come è fatto un ovulo, conoscere le strutture che governano posizione e orientamento del futuro embrione e, soprattutto, come fa il polline a penetrare nell’ovulo. È quello che fece Brown analizzando l’ovulo della Kingia. Fu lo studio di questa pianta (una angiosperma monocotiledone) che gli fornì la chiave per penetrare i misteri della gimnospermia.
Nel post (che segue un precedente articolo dove abbiamo raccontato come la scoperta della gimnospermia abbia influenzato i sistematici di metà Ottocento) cerchiamo di ricostruire il percorso intellettuale di Brown attraverso gli scritti originali. Un lavoro faticoso ma esaltante che ci farà rivivere un importantissimo momento della storia della botanica e nel contempo ci consentirà di apprendere molte cose sull’anatomia dell’ovulo, mostrando ancora una volta l’efficacia didattica della storia della scienza: non un racconto ma un modo per apprendere operativamente nozioni e concetti, e non già in maniera dichiarativa come troppo spesso avviene nei manuali.
1 – Descrizione dell’ovulo non fecondato di Kingia australis
(ovvero come è fatto l’ovulo delle angiosperme)
Il nome Kingia australis fu imposto da Brown in Annales des sciences naturelles, fascicolo 8, Parigi, 1826.

La pagina 211. Brown dichiara i suoi intenti: si parlerà della struttura dell’uovo non fecondato nelle piante a fiore (fanerogame) e del fiore femminile di cicadi e conifere.
Egli rivendica da subito la novità e originalità della ricerca in ragione, soprattutto, dei risultati ottenuti (vedi seconda parte) sulla posizione e orientamento che assumerà l’embrione.


La prima parte dell’articolo è dedicata alla descrizione della pianta (il protologo). Noi siamo interessati al solo ovulo.

Ovuli eretti, inseriti sul fondo dell’angolo interno del loculo poco sopra la propria base, obovati, compressi a forma di lente, privi di ali: la Testa [rivestimento esterno, oggi tegumento esterno o primina] è perforata alla sua stessa base, che è alquanto acuta, da un minuto forame [micropilo]; è presente una seconda membrana anch’essa perforata [oggi detta tegumento interno o secondina] [Brown ne specifica una peculiare caratteristica della quale però non ci occuperemo]; il nucleo [nucella] è conforme alla cavità della membrana, inserito alla base della stessa, per il resto libero, polposo e solido, con l’apice alquanto acuto e liscio che raggiunge l’apertura [il micropilo] della membrana interna.
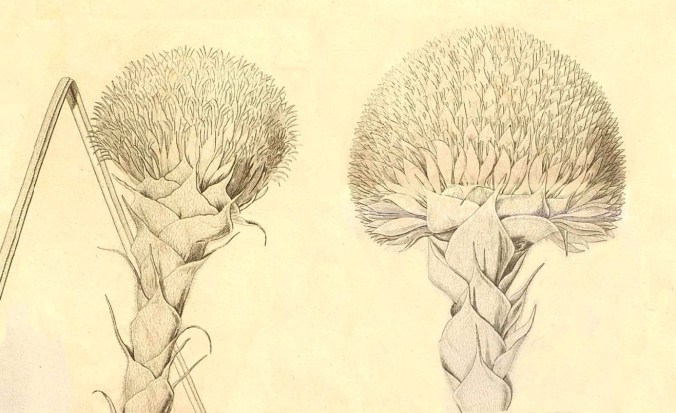
Esiste una comunicazione dal titolo Character and description of Kingia, alla Linnean Society of London, 1825 (un anno prima della pubblicazione sugli Annales) in cui compare una illustrazione della pianta.
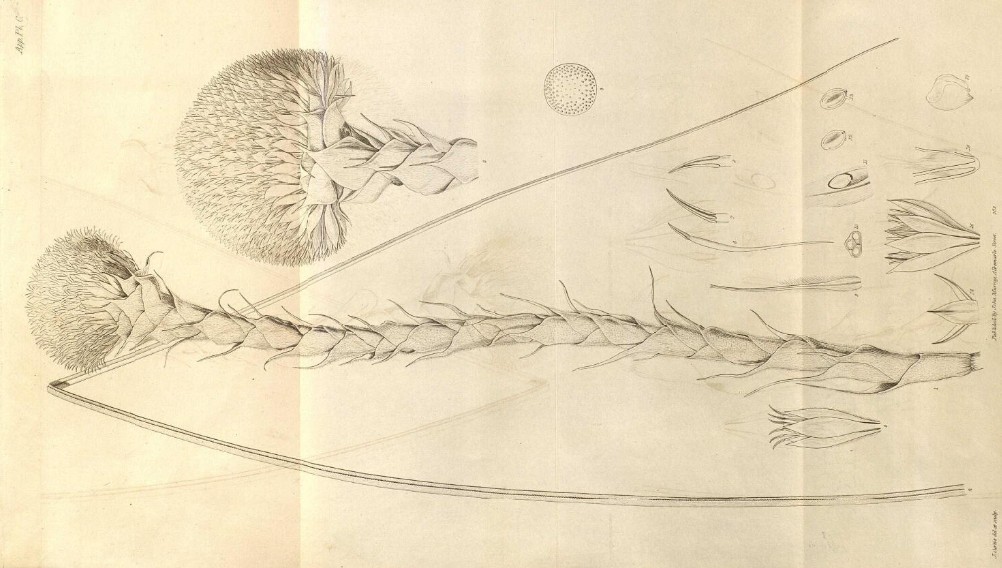
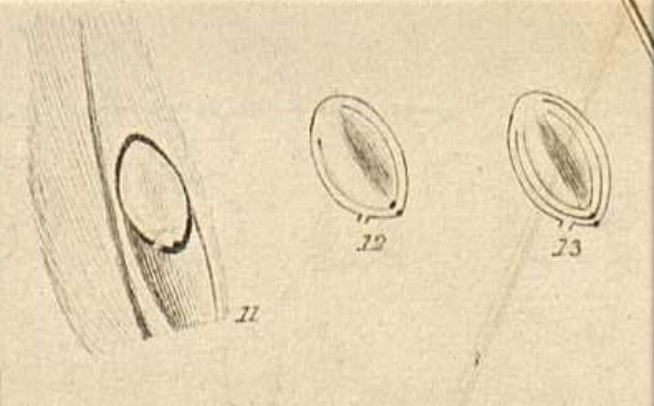
Questo è quello che Brown vedeva nel suo microscopio. (11) Particolare della sezione longitudinale dell’ovario con l’ovulo intero che occupa la loggia ed è attaccato alla placenta con un piedino (oggi lo chiamiamo funicolo). (12) Sezione dell’ovulo col funicolo, il rivestimento disegnato non in dettaglio, e due puntini allineati che rappresentano il micropilo. (13) In questa ulteriore sezione si distinguono i due tegumenti (primina all’esterno e secondina all’interno). L’interno della nucella è indifferenziato, Brown parla di un oggetto ‘polposo e solido’, ci vorranno strumenti molto più potenti per risolverlo. (Pagina 5)
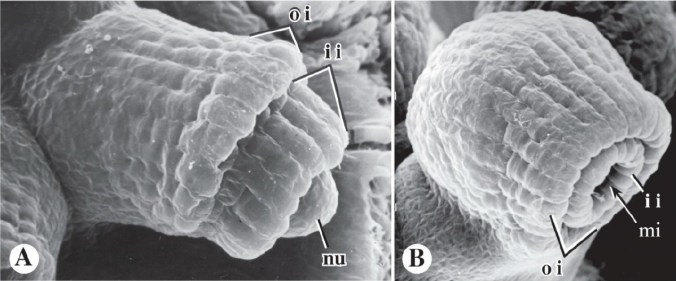
Urge una foto, e che foto: una scansione al microscopio elettronico (M. G. Simpson, Plant Systematics, Academic Press, 2010 – devo questa foto alle dispense di un corso di sistematica, intenso e affascinante, tenuto dal Professor Francesco Dal Grande – Unipd). Ovulo di Arabidopsis thaliana (una angiosperma molto studiata). A) stadio precoce, si distinguono la nucella (nu) e i due tegumenti interno (ii) e esterno (oi). Stanno crescendo dal basso verso l’alto. B) stadio maturo, una volta cresciuti i due tegumenti lasciano un buco, il micropilo (mi).
2 – Disposizione dell’embrione rispetto al micropilo
Nella seconda pare dell’articolo Brown enuncia una regola generale sulla disposizione che assumerà l’embrione.

La regola, già intuita nel lontano 1814, è subito enunciata: la radichetta (dell’embrione) si troverà sempre puntata verso il micropilo (l’ombelico lo chiama qui, in altri luoghi lo dice forame).
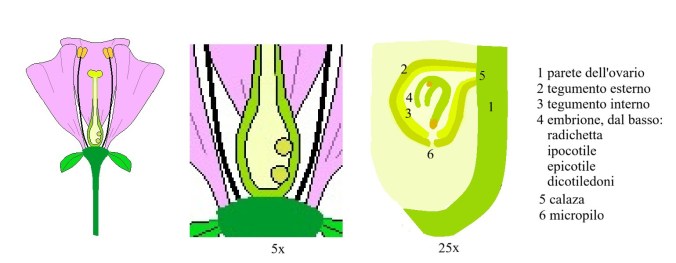
Successivi ingrandimenti. Brown lo ignorava, ma il polline non entra direttamente dal micropilo. Oggi sappiamo che il tubetto pollinico percorre la parete dell’ovario (1), penetra nel loculo dell’ovario, entra dal micropilo (6) e feconda l’ovulo (più esattamente la cellula uovo) dando luogo all’embrione nel quale, a maturazione completa, distinguiamo la radichetta e tutto il resto (4).

Questa seconda parte prosegue, e si conclude, con una lunga escursione storica nella quale Brown elenca gli scienziati che avevano affrontato l’argomento rivelandoci quanto fossero andati vicini alla sua scoperta o quanto se ne fossero allontanati. Si tratta di un modo di fare comune (forse ancora oggi) che serve sia a ribadire le tesi enunciate sia a marcare le differenze colle interpretazioni date in precedenza. Ad esempio nel caso della prima citazione Brown ci dice che Nehemiah Grew (1641-1712) osservò una ‘piccola apertura, posta di fronte alla radichetta dell’embrione’, ma che la interpretò come un foro di aerazione.
È sempre di grande interesse leggere queste cose: ci aiutano a capire come la Natura mostri a tutti le stesse cose, ma molte sono le interpretazioni che se ne possono dare. Vi invito a proseguire nella lettura. Per parte nostra proseguiamo con una nuova parte dell’articolo senza altri indugi.
3 – Il ruolo del micropilo nel denudare l’ovulo delle gimnosperme

La terza parte dell’articolo (pagina 232) ha titolo ‘Sulla struttura dell’uovo femminile nelle Cicadi e nelle Conifere’. Riflettendo sul ruolo del micropilo Brown ragiona così: che il polline entri dal micropilo è una buona ipotesi (dopotutto è in prossimità del buco che cresce l’embrione), ma se si trovassero piante con ovuli esposti non racchiusi in un ovario, l’ipotesi sarebbe altamente corroborata, perché resterebbe provato che l’ovario non è essenziale per la fecondazione la quale può avvenire anche in sua assenza. Ma queste piante esistono eccome, sono le cicadi, le conifere, le efedre e le Gnetum, insomma (a parte il ginkgo) tutte quelle che noi oggi sappiamo essere gimnosperme.

Egli continua ad argomentare: il fatto che non sia l’ovario bensì l’ovulo delle fanerogame a somigliare al così detto fiore di cicadi e conifere (noi diremmo delle gimnosperme) vuol dire che quello che noi abbiamo finora chiamato fiore di cicadi e conifere altro non è che un ovulo lui stesso. Ma la prova provata è che non solo c’è somiglianza esterna (tra fiore di cicadi e conifere e ovulo delle fanerogame) ma esiste corrispondenza nella struttura: in entrambi è presente il micropilo e la radichetta punta verso quest’ultimo. Così il cerchio si chiude.
Godiamoci ora una breve selezione di foto di ovuli nudi di gimnosperme; impressionante la varietà di soluzioni adottate dalla Natura.














Anche per questa terza parte il Nostro fa un excursus storico, ma noi non lo seguiamo, siamo invece interessati ad altri argomenti affrontati verso la fine dell’articolo.

Uno di questi (pagina 234) concerne il numero di tegumenti che racchiudono l’ovulo. Se ricordate nelle fanerogame erano due, Brown afferma che nelle cicadi e nella più parte delle conifere è uno solo. Ma i dubbi sono tanti poiché in altre specie di ‘non-fanerogame’ trova un numero di tegumenti variabile (oggi lo sappiamo per certo), così non si azzarda a trarre alcuna conclusione.

Brown continua a ragionare (pagina 240): se tra fanerogame da una parte e cicadi e conifere dall’altra, ci sono differenze nelle strutture riproduttive femminili, allora ‘ci si dovrebbe aspettare’ una differenza anche riguardo alle strutture maschili. Insomma alle antere delle angiosperme cosa corrisponde nelle gimnosperme? È una buona domanda alla quale però il buon Robert non se la sentirà di dare una risposta definitiva.

Strobilo maschile di Thuja orientalis, le sacche polliniche sono attaccate a peduncoli sottili ortogonali all’asse centrale, sono protette da brattee a cupola. Struttura, col senno di poi, molto diversa da un amento di angiosperma.

A modo di conclusione
Anche se nell’articolo molto resta in sospeso, siamo ugualmente ammirati dal numero e dalla qualità di argomenti prodotti da tale mente effervescente. Essi costituiranno una eredità per gli scienziati successivi, alcuni dei quali capirono subito l’importanza della scoperta (ne sono testimoni le numerose traduzioni dell’articolo) e si misero alacremente al lavoro. Tuttavia è doveroso dire che la scoperta richiese molto tempo prima di essere riconosciuta e assimilata. Non ci stupiamo, noi che, pur istruiti dai moderni manuali, abbiamo così tanto faticato a seguire le argomentazioni prodotte. Il primo sistematico che introdusse la scoperta in una tassonomia fu, come abbiamo visto in altro post, Brongniart; e possiamo considerare indicativo delle difficoltà dell’argomento il tentativo maldestro di Lidley raccontato nello stesso articolo. Ma bisognerà attendere Eichler per vedere messi in bell’ordine tutti gli attori di questa storia straordinaria: vi rimando all’indice del suo Bluthendiagramme. Un libro, per altri motivi, ben noto a questo blog.
